양봉 꿀벌의 경쟁 서식지에서의 미포자충 기생충의 뚜렷한 독성
페이지 정보

본문
꿀벌의 경쟁 서식지에서의 미포자충 기생충의 뚜렷한 독성
자연 생태계에서 기생충은 종종 여러 숙주 종을 감염시키는데, 특히 숙주가 서식지를 공유할 때, 숙주 간 전파를 용이하게 하고 전통적인 숙주-기생충 공진화 역학을 변화시킨다. 이 연구에서는 동부 꿀벌( Apis cerana )과 서부 꿀벌( Apis mellifera )의 미포자충 기생충인 Nosema ceranae를 조사하여 독성과 증식 역학을 평가한다. 접종 실험을 사용하여 꿀벌 사망률과 기생충 포자 부하를 측정하여 독성과 증식을 유추했다. 또한 꿀벌과 기생충 모두에 대한 시계열 전사체 분석은 숙주-병원체 상호 작용에 대한 통찰력을 제공한다. 결과에 따르면 N. ceranae는 A. mellifera 에서 낮은 사망률로 더 많은 포자를 생성 하지만 A. cerana 에서는 낮은 포자 생성으로 더 높은 사망률을 유발한다 . 이 기생충은 또한 숙주 유전자 발현을 억제하는데, A. cerana 에서 더 강한 억제가 관찰되었다 . 이러한 발견은 N. ceranae가 A. mellifera 에서 낮은 독력과 높은 증식에 적응했지만 A. cerana 에서는 높은 독력과 제한된 증식을 보인다는 것을 시사합니다 . 이 연구는 다중 숙주 시스템에서 독력과 증식 간의 뚜렷한 상충 관계의 진화를 강조하여 기생충-숙주 역학과 그 생태적 의미에 대한 귀중한 통찰력을 제공합니다.
1 서론
숙주 사망률은 일반적으로 기생충의 독성을 평가하는데, 이는 정적이지 않고 숙주 생태에 따라 진화합니다( Gowler et al., 2023 ). 대부분의 진화적 독성 이론은 기생충과 숙주 적합도 간의 상충 관계를 연결합니다. 예를 들어, 기생충의 높은 증식은 다른 개체로의 전파를 증가시킵니다. 그러나 높은 증식은 숙주를 빠르게 죽이고 기생충 전파를 줄일 수 있습니다( de Roode et al., 2008 ). 따라서 공진화된 숙주-기생충에서 균형 잡힌 전파와 독성이 예상되었습니다( Acevedo et al., 2019 ).
자연 생태계에서 기생충은 이동하여 대체 숙주 생물을 탐색합니다.숙주 종의 계통 발생적 관계와 근접성은 숙주 이동의 성공에 영향을 미치며, 밀접한 관련 종이 기생충을 공유할 가능성이 가장 높습니다( Engelstädter and Fortuna, 2019 ). 예를 들어, Nosema ceranae는 아시아 꿀벌 Apis cerana 와 유럽 꿀벌 Apis mellifera를 모두 감염시킵니다 ( Fries et al., 1996 ; Higes et al., 2007 ). 감염은 포자에 오염된 꽃꿀을 섭취하는 것으로 시작됩니다. 포자는 중장 루멘에서 발아하고 포자질을 극관을 통해 상피 세포로 주입합니다( Gisder et al., 2011 ). 감염된 꿀벌은 억제된 세포사멸( Higes et al., 2013 ), 미성숙한 노화( Paris et al., 2018 ), 단축된 수명( Eiri et al., 2015 ), 손상된 비행( Gage et al., 2018 )을 보입니다.
아시아에서 두 꿀벌 종은 서식지와 은신처 자원을 놓고 경쟁하며 기생충인 N. ceranae 의 유행은 두 꿀벌 종 모두에서 70%가 넘었습니다( Li et al., 2012 ; Yang et al., 2013 ; Jack et al., 2016 ). 따라서 기생충이 두 꿀벌 종 사이를 오갈 가능성이 높습니다( Graystock et al., 2015 ). N. ceranae 감염은 유럽과 아시아 꿀벌 모두에서 전역 유전자 발현을 변화시킵니다( Holt et al., 2013 ; Fan et al., 2022 ). 기생충 감염은 유럽 꿀벌에서 군집 실패를 유발합니다( Higes et al., 2008 ; Botías et al., 2013 ). 비교적으로 아시아 꿀벌에서의 독성은 불분명합니다. 이전에 우리는 숙주 서식지 공유가 기생충 유전자 흐름을 증가시킨다는 것을 발견했습니다( Ke et al., 2022 ). 이 후속 연구에서 우리는 두 종류의 꿀벌 종과 미포자충 기생충을 사용하여 숙주 서식지 공유가 기생충 독성을 어떻게 형성하는지 조사했습니다. 우리는 공유 서식지에서 밀접한 관련이 있는 두 꿀벌에서 독성과 증식 사이에 뚜렷한 상충 관계를 발견했습니다.
2 재료 및 방법
2.1 윤리적 성명
꿀벌 Apis mellifera 와 Apis cerana는 보호종도 멸종위기종도 아닙니다. 이 연구에는 윤리적 승인이 필요하지 않습니다.
2.2 숙주 및 기생충 공급원
우리는 두 종류의 꿀벌에서 기생충의 독성과 증식을 연구하기 위해 2x2 요인 실험을 설계했습니다. 우리는 두 가지 기생충 공급원인 P Acer (꿀벌 Apis cerana 에서 정제된 기생충 포자 )와 P Amel (꿀벌 Apis mellifera 에서 정제된 기생충 포자 )과 두 가지 숙주 종인 H Acer (숙주 꿀벌 Apis cerana )와 H Amel (숙주 꿀벌 Apis mellifera )을 사용했습니다( 보충 표 S1 ). 우리는 접종을 위해 다른 벌집의 벌을 합쳤고, 이 다중 숙주 기생충 실험을 위한 대조군으로 감염되지 않은 벌(H Acer 와 H Amel )을 합쳤습니다. 꿀벌 군집은 장시 농업 대학의 실험 양봉장에서 유지됩니다.
2.3 기생충 분리, 접종 및 RNA 추출
A. mellifera 와 A. cerana 각각의 꿀벌 300마리를 곤충 그물을 사용하여 벌집 입구 근처에서 포획했습니다. 중장을 절개하여 균질화하여 N. ceranae 포자를 분리한 다음 Percoll 농도 구배( Chen et al., 2013 )를 사용하여 추가로 정제했습니다. 포자는 혈구계를 사용하여 광학 현미경으로 세었습니다. H Amel 과 H Acer 의 세 벌집에서 밀봉된 육아 틀을 인큐베이터에 보관하여 새로 부화한 벌을 모았습니다(35°C, 습도 75%).
새로 부화한 꿀벌 일벌(H Acer 및 H Amel )은 포자 10 5 가 포함된 설탕 용액 2 µL를 개별적으로 접종했습니다 . 새로 부화한 꿀벌(부화 후 < 24시간)은 감염되지 않은 대조군으로 포자가 없는 설탕 용액 2 µL를 각각 먹였습니다. 각 처리군에서 150마리의 꿀벌에게 접종하고, 집단을 인큐베이터(35°C, 습도 75%)에서 3개의 사육 컵(컵당 벌 50마리)으로 나누었습니다(3회 반복, 보충 표 S1 ). 일반적인 벌 반응과 기생충 증식을 조사하기 위해 말입니다. 실험하는 동안, 자당(50% w/w)을 유일한 음식으로 자유롭게 제공했습니다 . 각 사육 컵에서 RNA-seq를 위해 1~5 dpi(접종 후 일)까지 24시간 간격으로 벌 3마리를 수집했습니다. 남은 벌들은 14dpi에서 포자를 세기 위해 해부되었습니다.
2.4 RNA 추출 및 라이브러리 제조
기생충이 중장의 상피 세포를 감염시키므로, RNA-seq를 위해 중장 조직을 절개했습니다. 하루에 컵당 3마리의 벌을 절개하여 Trizol을 사용하여 RNA 추출을 위해 모았습니다. 라이브러리는 NEBNext Ultra RNA 키트를 사용하여 준비했습니다. 총 90개의 RNA 라이브러리(5일 * 6회 처리 * 3회 반복)를 Illumina NovaSeq 6000 플랫폼에서 시퀀싱했습니다.
2.5 생물정보학 및 통계학
RNA 판독(150bp, paired-end)의 품질은 Fastqc( Andrews, 2010 )를 사용하여 먼저 확인했고 기본 매개변수가 있는 Seqtk 패키지( Li, 2022 )를 사용하여 트리밍했습니다. 처리된 판독은 기본 매개변수가 있는 Hisat2 패키지( Kim et al., 2015 ; Diao et al., 2018; Wallberg et al., 2019 ; Huang et al., 2021)를 사용하여 각각 N. ceranae (버전 Ncer 3.0), A. cerana (버전 CC1.0 ) 및 A. mellifera( 버전 Hav3.1 ) 유전체에 정렬했습니다. 유전자당 판독 횟수는 bedtools( Quinlan and Hall, 2010 )를 사용하여 검색했습니다. 그룹 내 분산은 edgeR 패키지를 사용하여 유의하게 조절된 유전자를 결정하고 FDR(거짓 발견율)에 대해 조정하기 위해 3회 반복에서 계산했습니다( Robinson et al., 2010 ). FDR < 0.05인 유전자는 유의하게 조절된 유전자로 정의되었습니다. 유전자 온톨로지(GO) 용어는 EggNOG-mapper를 사용하여 검색했고, 풍부화 분석은 조정된 가중 ks 검정이 있는 TopGo 패키지를 사용하여 수행했습니다( Robinson et al., 2010 ; Alexa and Rahnenfuhrer, 2021 ). 꿀벌 생존은 FDR에 대해 조정된 생존 패키지의 Kaplan-Meier 추정치를 사용하여 분석했습니다( R Core Team, 2013 ; Therneau, 2022 ). 처리 그룹 간 포자 부하의 분산은 Wilcoxon 순위 검정으로 분석했고, p 값은 거짓 양성을 줄이기 위해 FDR로 조정했습니다. 포자원과 요일이 유전자 발현에 미치는 영향은 ANONA를 사용하여 분석했습니다. 여기서 요일과 기생충원은 고정 요인이고 반복은 임의 요인이었습니다. 상향 및 하향 조절된 유전자의 수는 피어슨 카이 제곱 검정을 사용하여 분석했습니다.
3개의 결과
3.1 기생충은 꿀벌에서 뚜렷한 독성을 보입니다.
감염되지 않은 H Amel 은 가장 높은 생존율(99.4% 생존율)을 보였고, 그 다음으로 P Acer 에 감염된 H Amel (H Amel _P Acer , 생존율 98.5%)과 H Amel _P Amel 그룹(생존율 96.9%) 순이었다. H Acer _P Acer 그룹 의 벌은 가장 낮은 생존율(생존율 86.6%)을 보였다. 기생충 출처(P Amel 및 P Acer )는 H Amel 의 생존에 미미한 영향을 미치는 것으로 나타났다 (H Amel _P Acer 대 H Amel _P Amel , Kaplan-Meier 검정, P > 0.05)( 그림 1A , 보충 표 S2 ). 비교적 기생충 P Acer은 P Amel 보다 H Acer 에서 유의하게 높은 사망률을 유발한다 (Coxph 검정, P < 0.001). 감염되지 않은 H Amel 은 감염되지 않은 H Acer 보다 훨씬 더 잘 생존한다 (Coxph 검정, P < 0.01). 전반적으로 기생충은 H Amel 보다 H Acer 에서 더 높은 사망률을 유발합니다. 또한 감염되지 않은 H Amel 과 감염되지 않은 H Acer를 사용하여 두 꿀벌 종 간의 사망률 분산을 정규화합니다 . 다시 말하지만, 기생충은 H Amel 보다 H Acer 에서 더 높은 사망률을 유발합니다 ( P < 0.001, 보충 그림 S1 ).
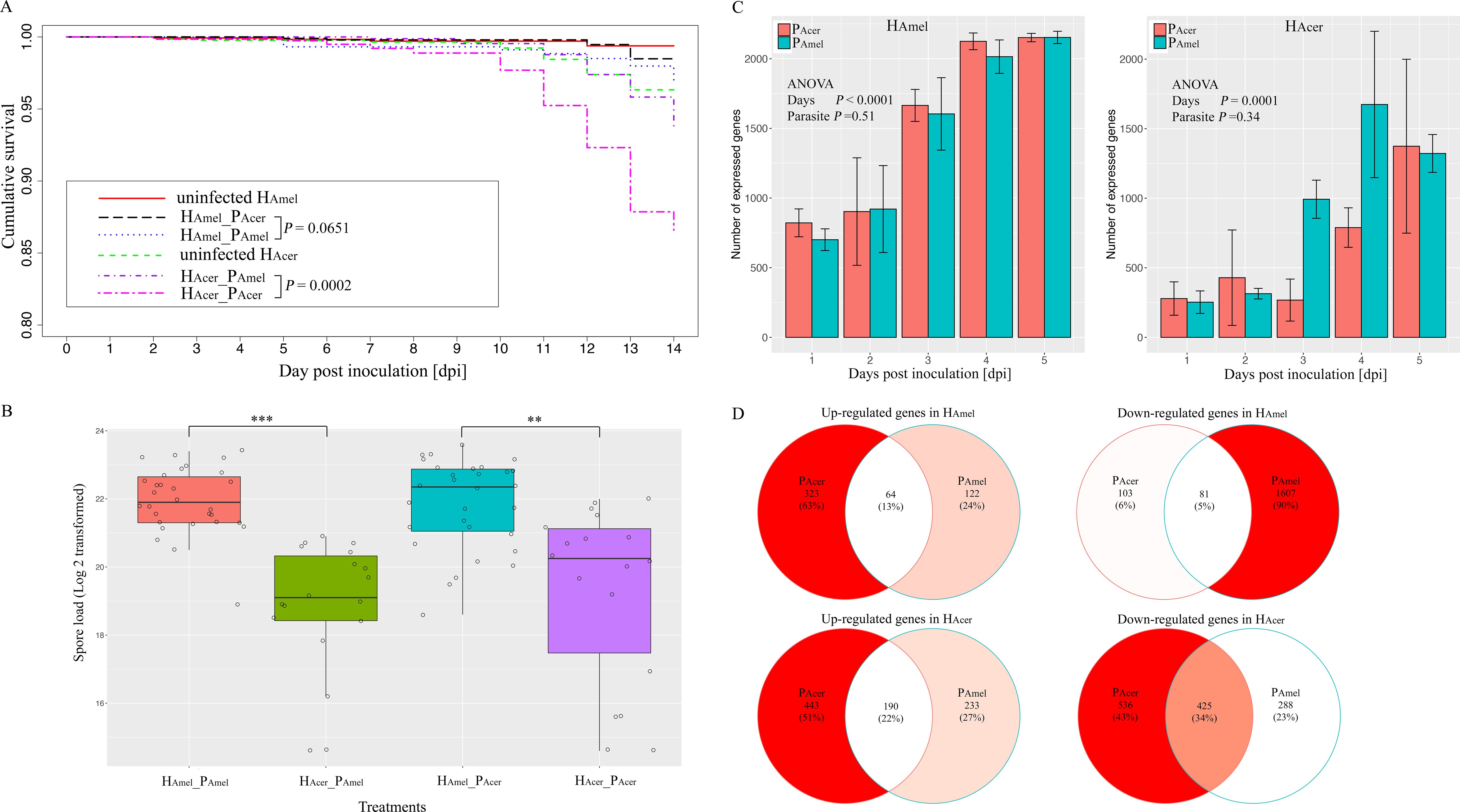
그림 1. 2개 기생충과 2개 숙주 시스템에서 기생충의 전파 및 독성과 기생충에 대한 벌의 반응. (A) 꿀벌의 누적 생존. 벌의 생존에 대한 기생충 출처의 영향은 미미했습니다. P Acer는 H Acer 에서 P Amel 보다 상당히 높은 사망률을 초래했습니다 . (B) 두 꿀벌 종에서의 기생충 증식 변이. 기생충은 H Acer 보다 H Amel 에서 더 많은 포자를 생성했습니다 . 또한, 포자 부하에 대한 기생충 출처의 영향은 두 숙주 모두에서 미미했습니다. (C) 두 숙주에서 발현된 기생충 유전자의 수. 기생충 출처에 관계없이 A . cerana 에서 A . mellifera 보다 적은 기생충 유전자가 발현되었습니다. (D) 두 기생충 분리체에 반응하는 공유 및 고유한 조절 숙주 유전자의 벤 다이어그램. 전반적으로 벌 유전자는 감염에 의해 하향 조절되었고, 유전자 하위 집합은 두 기생충 출처 모두에 반응했습니다. **는 P < 0.01 에서의 유의수준을 나타냅니다 . ***는 P < 0.001 에서의 유의수준을 나타냅니다 . 오차 막대는 표준편차를 나타냅니다.
3.2 기생충은 상당한 증식 변이를 보입니다.
감염되지 않은 벌에서는 포자가 발견되지 않았습니다.포자 부하량은 감염된 네 꿀벌 그룹에 고르게 분포되지 않았습니다(Kruskal-Wallis 검정, χ2 = 43.1, df=3, P < 0.0001, 그림 1B ).기생충은 P Amel 또는 P Acer 에 감염되었을 때 H Amel 에서 H Acer 보다 더 많은 포자를 생산합니다 (Wilcoxon 순위 합계 검정, df=1, P < 0.001)( 보충 표 S3 ).따라서 숙주 종은 기생충 증식에 상당한 영향을 미칩니다(F=32.3, df=2, ANOVA, P < 0.0001).
3.3 H Amel 에서 H Acer 보다 더 높은 기생충 유전자 발현 프로필
기생충 유전자 발현 프로필을 조사하기 위해 두 숙주 종에서 기생충 전사체를 정량화합니다( 그림 1C ). H Acer는 H Amel 보다 기생충 유전자 발현을 억제하는 경향을 더 강하게 보입니다 (F=77.1, df=1, ANOVA, P < 0.0001). H Amel을 감염시켰을 때 P Amel 에서 P Acer 보다 유의하게 적은 수의 상향 조절된 유전자가 관찰되었습니다 (피어슨 카이 제곱 검정, χ 2 = 20.8, df=2, P < 0.0001). 가장 높은 분산은 4 dpi에서 나타났고, H Amel에서 상향 조절된 유전자의 수는 P Acer(36개 유전자)에서 P Amel(12개 유전자)보다 3배 더 높습니다 ( 보충 표 S4 ) . 따라서 기생충은 H Amel 에서 H Acer 보다 더 많은 수의 유전자와 전사 수준을 발현합니다 .
관련링크
댓글목록
등록된 댓글이 없습니다.
